Что такое клеточная мембрана в биологии определение. Строение клеточной мембраны
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии - гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны - молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны - цепочки олигосахаридов , формирующие гликокаликс
Биологическая мембрана включает и различные белки : интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов .
Функции биомембран
- барьерная - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов . Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная - через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза .
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия , при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза , которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная - обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая - обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных - межклеточное вещество.
- энергетическая - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная - некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная - мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса .
- маркировка клетки - на мембране есть антигены, действующие как маркеры - «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды , гликолипиды и холестерол . Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп ) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы , отделённые от гиалоплазмы мембранами . К одномембранным органеллам относятся эндоплазматическая сеть , аппарат Гольджи , лизосомы , вакуоли , пероксисомы ; к двумембранным - ядро , митохондрии , пластиды . Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза , аминокислоты , жирные кислоты , глицерол и ионы , причем сами мембраны в известной мере активно регулируют этот процесс - одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клеки наружу: диффузия , осмос , активный транспорт и экзо- или эндоцитоз . Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних - активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами - интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход . Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия .
Ссылки
- Bruce Alberts, et al. Molecular Biology Of The Cell . - 5th ed. - New York: Garland Science, 2007. - ISBN 0-8153-3218-1 - учебник по молекулярной биологии на англ. языке
- Рубин А.Б. Биофизика, учебник в 2 тт. . - 3-е издание, исправленное и дополненное. - Москва: издательство Московского университета, 2004. - ISBN 5-211-06109-8
- Геннис Р. Биомембраны. Молекулярная структура и функции: перевод с англ. = Biomembranes. Molecular structure and function (by Robert B. Gennis). - 1-е издание. - Москва: Мир, 1997. - ISBN 5-03-002419-0
- Иванов В.Г., Берестовский Т.Н. Липидный бислой биологических мембран. - Москва: Наука, 1982.
- Антонов В.Ф., Смирнова Е.Н., Шевченко Е.В. Липидные мембраны при фазовых переходах. - Москва: Наука, 1994.
См. также
- Владимиров Ю. А., Повреждение компонентов биологических мембран при патологических процессах
Wikimedia Foundation . 2010 .
Биологические мембраны - общее название функционально активных поверхностных структур, ограничивающих клетки (клеточные, или плазматические мембраны) и внутриклеточные органеллы (мембраны митохондрий, ядер, лизосом, эндоплазматического ретикулума и др.). Они содержат в своем составе липиды, белки,гетерогенные молекулы (гликопротеины,гликолипиды)и в зависимости от выполняемой функции многочисленные минорные компоненты: коферменты, нуклеиновые кислоты, антиоксиданты, каротиноиды, неорганические ионы и т. п.
Согласованное функционирование мембранных систем – рецепторов, ферментов, транспортных механизмов - помогает поддерживать гомеостаз клетки и в то же время быстро реагировать на изменения внешней среды.
К основным функциям биологических мембран можно отнести:
· отделение клетки от окружающей среды и формирование внутриклеточных компартментов (отсеков);
· контроль и регулирование транспорта огромного многообразия веществ через мембраны;
· участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
· преобразование энергии пищевых органических веществ в энергию химических связей молекул АТФ.
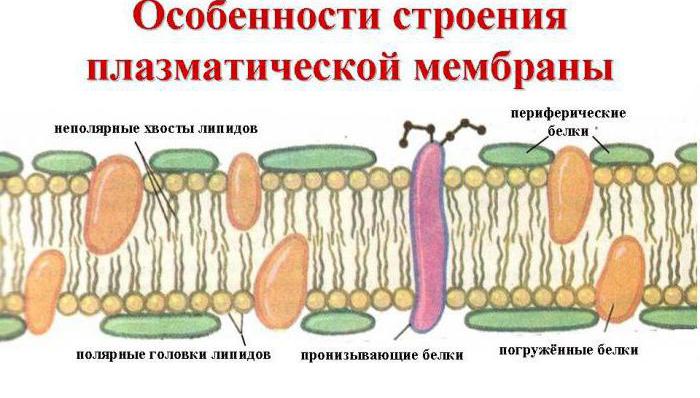
Молекулярная организация плазматической (клеточной) мембраны у всех клеток примерно одинакова: она состоит из двух слоев липидных молекул с множеством включенных в нее специфических белков. Одни мембранные белки обладают ферментативной активностью, тогда как другие связывают питательные вещества из окружающей среды и обеспечивают их перенос в клетку через мембраны. Мембранные белки различают по характеру связи с мембранными структурами. Одни белки, называемые внешними или периферическими , непрочно связаны с поверхностью мембраны, другие, называемые внутренними или интегральными , погружены внутрь мембраны. Периферические белки легко экстрагируются, тогда как интегральные белки могут быть выделены только при помощи детергенов или органических растворителей. На рис. 4 представлена структура плазматической мембраны.
Внешние, или плазматические, мембраны многих клеток, а также мембраны внутриклеточных органелл, например, митохондрий, хлоропластов удалось выделить в свободном виде и изучить их молекулярный состав. Во всех мембранах имеются полярные липиды в количестве, составляющем в зависимости от типа мембран от 20 до 80% ее массы, остальное приходится главным образом на долю белков. Так, в плазматических мембранах животных клеток количество белков и липидов, как правило, примерно одинаково; во внутренней митохондриальной мембране содержится около 80% белков и только 20% липидов, а в миелиновых мембранах клеток мозга наоборот, около 80% липидов и только 20% белков.
Рис. 4. Структура плазматической мембраны
Липидная часть мембран представляет собой смесь различного рода полярных липидов. Полярные липиды, к числу которых относятся фосфоглицеролипиды, сфинголипиды, гликолипиды не запасаются в жировых клетках, а встраиваются в клеточные мембраны, причем в строго определенных соотношениях.
Все полярные липиды в мембранах постоянно обновляются в процессе метаболизма, при нормальных условиях в клетке устанавливается динамическое стационарное состояние, при котором скорость синтеза липидов равна скорости их распада.
В мембранах животных клеток присутствуют в основном фосфоглицеролипиды и в меньшей степени сфинголипиды; триацилглицеролы обнаруживаются лишь в следовых количествах. Некоторые мембраны животных клеток, в особенности наружная плазматическая мембрана, содержит значительные количества холестерола и его эфиров (рис.5).

Рис.5. Мембранные липиды
В настоящее время общепринятой моделью строения мембран является жидкостно-мозаичная, предложенная в 1972 году С. Синджером и Дж. Николсоном.
Согласно ей белки можно уподобить айсбергам, плавающим в липидном море. Как уже указывалось выше, существуют 2 типа мембранных белков: интегральные и периферические. Интегральные белки пронизывают мембрану насквозь, они являются амфипатическими молекулами . Периферические белки не пронизывают мембрану и связаны с ней менее прочно. Основной непрерывной частью мембраны, то есть ее матриксом, служит полярный липидный бислой. При обычной для клетки температуре матрикс находится в жидком состоянии, что обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами в гидрофобных хвостах полярных липидов.
Жидкостно-мозаичная модель предполагает также, что на поверхности расположенных в мембране интегральных белков имеются R-группы аминокислотных остатков (в основном гидрофобные группы, за счет которых белки как бы «растворяются» в центральной гидрофобной части бислоя). В то же время, на поверхности периферических, или внешних белков, имеются в основном гидрофильные R-группы, которые притягиваются к гидрофильным заряженным полярным головкам липидов за счет электростатических сил. Интегральные белки, а к ним относятся ферменты и транспортные белки, обладают активностью только в том случае, если находятся внутри гидрофобной части бислоя, где они приобретают необходимую для проявления активности пространственную конфигурацию (рис.6). Следует еще раз подчеркнуть, что ни между молекулами в бислое, ни между белками и липидами бислоя не образуется ковалентных связей.

Рис.6. Мембранные белки
Мембранные белки могут свободно перемещаться в латериальной плоскости. Периферические белки буквально плавают на поверхности бислойного «моря», а интегральные белки, подобно айсбергам, почти полностью погружены в углеводородный слой.
В большинстве своем мембраны ассиметричны, то есть имеют неравноценные стороны. Эта ассиметричность проявляется в следующем:
· во-первых, в том, что внутренняя и внешняя стороны плазматических мембран бактериальных и животных клеток различаются по составу полярных липидов. Так, например, внутренний липидный слой мембран эритроцитов человека содержит в основном фосфатидилэтаноламин и фосфатидилсерин, а внешний – фосфатидилхолин и сфингомиелин.
· во-вторых, некоторые транспортные системы в мембранах действуют только в одном направлении. Например, в мембранах эритроцитов имеется транспортная система («насос»), перекачивающая ионы Nа + из клетки в окружающую среду, а ионы К + - внутрь клетки за счет энергии, освобождающейся при гидролизе АТФ.
· в-третьих, на внешней поверхности плазматических мембран содержится очень большое число олигосахаридных группировок, представляющих собой головки гликолипидов и олигосахаридные боковые цепи гликопротеинов, тогда как на внутренней поверхности плазматической мембраны олигосахаридных группировок практически нет.
Ассиметричность биологических мембрам сохраняется за счет того, что перенос индивидуальных молекул фосфолипидов с одной стороны липидного бислоя на другую очень затруднен по энергетическим соображениям. Полярная молекула липида способна свободно перемещаться на своей стороне бислоя, но ограничена в возможности перескочить на другую сторону.
Подвижность липидов зависит от относительного содержания и типа присутствующих ненасыщенных жирных кислот. Углеводородная природа жирнокислотных цепей сообщает мембране свойства текучести, подвижности. В присутствии цис-ненасыщенных жирных кислот силы сцепления между цепями слабее, чем в случае одних насыщенных жирных кислот, и липиды сохраняют высокую подвижность и при низкой температуре.
На внешней стороне мембран имеются специфические распознающие участки, функция которых состоит в распознавании определенных молекулярных сигналов. Например, именно посредством мембраны некоторые бактерии воспринимают незначительные изменения концентрации питательного вещества, что стимулирует их движение к источнику пищи; это явление носит название хемотаксиса .
Мембраны различных клеток и внутриклеточных органелл обладают определенной специфичностью, обусловленной их строением, химическим составом и функциями. Выделяют следующие основные группы мембран у эукариотических организмов:
· плазматическая мембрана (наружная клеточная мембрана, плазмалемма),
· ядерная мембрана,
· эндоплазматический ретикулум,
· мембраны аппарата Гольджи, митохондрий, хлорпластов, миелиновых оболочек,
· возбудимые мембраны.
У прокариотических организмов помимо плазматической мембраны существуют внутрицитоплазматические мембранные образования, у гетеротрофных прокариот они называются мезосомами. Последние образуются впячиванием внуть наружной клеточной мембраны и в некоторых случаях сохраняют с ней связь.
Мембрана эритроцитов состоит из белков (50%), липидов (40%) и углеводов (10%). Основная часть углеводов (93%) связана с белками, остальная – с липидами. В мембране липиды расположены асимметрично в отличие от симметричного расположения в мицеллах. Например, кефалин находится преимущественно во внутреннем слое липидов. Такая асимметрия поддерживается, по-видимому, за счет поперечного перемещения фосфолипидов в мембране, осуществляемого с помощью мембранных белков и за счет энергии метаболизма. Во внутреннем слое эритроцитарной мембраны находятся в основном сфингомиелин, фосфатидилэтаноламин, фосфатидилсерин, в наружном слое – фосфатидилхолин. Мембрана эритроцитов содержит интегральный гликопротеин гликофорин , состоящий из 131 аминокислотного остатка и пронизывающий мембрану, и так называемый белок полосы 3, состоящий из 900 аминокислотных остатков. Углеводные компоненты гликофорина выполняют рецепторную функцию для вирусов гриппа, фитогемагглютининов, ряда гормонов. В эритроцитарной мембране обнаружен и другой интегральный белок, содержащий мало углеводов и пронизывающий мембрану. Его называют туннельным белком (компонент а), так как предполагают, что он образует канал для анионов. Периферическим белком, связанным с внутренней стороной эритроцитарной мембраны, является спектрин.
Миелиновые мембраны , окружающие аксоны нейронов, многослойны, в них присутствует большое количество липидов (около 80%, половина из них – фосфолипидов). Белки этих мембран важны для фиксации лежащих друг над другом мембранных солев.
Мембраны хлоропластов . Хлоропласты покрыты двухслойной мембраной. Наружная мембрана имеет некоторое сходство с таковой у митохондрий. Помимо этой поверхностной мембраны в хлоропластах имеется внутренняя мембранная система – ламеллы . Ламеллы образуют или уплощенные пузырьки – тилакоиды, которые, располагаясь друг над другом, собираются в пачки (граны) или формируют мембранную систему стромы (ламеллы стромы). Ламеллы гран и стромы наружной стороне мембраны тилакоидов сосредоточены гидрофильные группировки, галакто- и сульфолипидов. Фитольная часть молекулы хлорофилла погружена в глобулу и находится в контакте в гидрофобными группами белков и липидов. Порфириновые ядра хлорофилла в основном локализованы между соприкасающимися мембранами тилакоидов гран.
Внутренняя (цитоплазматическая) мембрана бактерий по структуре сходна с внутренними мембранами хлоропластов и митохондрий. В ней локализованы ферменты дыхательной цепи, активного транспорта; ферменты, участвующие в образовании компонентов мембраны. Преобладающим компонентом бактериальных мембран являются белки: соотношение белок/липид (по массе) равно 3:1. Наружная мембрана грамотрицательных бактерий по сравнению с цитоплазматической содержит меньшее количество различных фосфолипидов и белков. Обе мембраны различаются по липидному составу. Во внешней мембране находятся белки, образующие поры для проникновения многих низкомолекулярных веществ. Характерным компонентом наружной мембраны является также специфический липополисахарид. Ряд белков наружной мембраны служит рецепторами для фагов.
Мембрана вирусов. Среди вирусов мембранные структуры характерны для содержащих нуклеокапсид, который состоит из белка и нуклеиновой кислоты. Это «ядро» вирусов окружено мембраной (оболочка). Она также состоит из двойного слоя липидов с включенными в него гликопротеинами, расположенными в основном на поверхности мембраны. У ряда вирусов (микровирусы) в мембраны входит 70-80% всех белков, остальные белки содержатся в нуклеокапсиде.
Таким образом, мембраны клеток представляют собой очень сложные структуры; составляющие их молекулярные комплексы образуют упорядоченную двумерную мозаику, что придает поверхности мембран биологическую специфичность.
Изучением строения организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.
Какие функции выполняет наружная клеточная мембрана
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция - это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.

Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать для построения свой защитной оболочки.
А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
Главные химические соединения, которые её образуют - это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.

Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы - структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.

Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы - распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.

Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.

Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.
Наружная клеточная мембрана (плазмалемма, цитолемма, плазматическая мембрана) животных клеток покрыта снаружи (т.е. на стороне, не контактирующей с цитоплазмой) слоем олигосахаридных цепей, ковалентно присоединенных к мембранным белкам (гликопротеины) и в меньшей степени к липидам (гликолипиды). Это углеводное покрытие мембраны называется гликокаликсом. Назначение гликокаликса пока не очень ясно; есть предположение, что эта структура принимает участие в процессах межклеточного узнавания.
У растительных клеток поверх наружной клеточной мембраны располагается плотный целлюлозный слой с порами, через которые осуществляется связь между соседними клетками посредством цитоплазматических мостиков.
У клеток грибов поверх плазмалеммы – плотный слой хитина .
У бактерий – муреина .
Свойства биологических мембран
1. Способность к самосборке после разрушающих воздействий. Это свойство определяется физико-химическими особенностями фосфолипидных молекул, которые в водном растворе собираются вместе так, что гидрофильные концы молекул разворачиваются наружу, а гидрофобные - внутрь. В уже готовые фосфолипидные слои могут встраиваться белки. Способность к самосборке имеет важное значение на клеточном уровне.
2. Полупроницаемость (избирательность в пропускании ионов и молекул). Обеспечивает поддержание постоянства ионного и молекулярного состава в клетке.
3. Текучесть мембран . Мембраны не являются жесткими структурами, они постоянно флюктуируют за счет вращательных и колебательных движений молекул липидов и белков. Это обеспечивает большую скорость протекания ферментативных и других химических процессов в мембранах.
4. Фрагменты мембран не имеют свободных концов , так как замыкаются в пузырьки.
Функции наружной клеточной мембраны (плазмалеммы)
Основными функциями плазмалеммы являются следующие: 1) барьерная, 2) рецепторная, 3) обменная, 4)транспортная.
1. Барьерная функция. Она выражается в том, что плазмалемма ограничивает содержимое клетки, отделяя его от внешней среды, а внутриклеточные мембраны разделяют цитоплазму на отдельные реакционные отсеки-компартменты .
2. Рецепторная функция. Одной из важнейших функций плазмалеммы является обеспечение коммуникации (связи) клетки с внешней средой посредством присутствующего в мембранах рецепторного аппарата, имеющего белковую или гликопротеиновую природу. Основная функция рецепторных образований плазмалеммы - распознавание внешних сигналов, благодаря которым клетки правильно ориентируются и образуют ткани в процессе дифференцировки. С рецепторной функцией связана деятельность различных регуляторных систем, а также формирование иммунного ответа.
Обменная функция определяется содержанием в биологических мембранах ферментных белков, являющихся биологическими катализаторами. Их активность меняется в зависимости от рН среды, температуры, давления, от концентрации как субстрата, так и самого фермента. Ферменты определяют интенсивность ключевых реакций метаболизма, а также их направленность.
Транспортная функция мембран. Мембрана обеспечивает избирательное проникновение в клетку и из клетки в окружающую среду различных химических веществ. Транспорт веществ необходим для поддержания в клетке соответствующего рН, надлежащей ионной концентрации, что обеспечивает эффективность работы клеточных ферментов. Транспорт поставляет питательные вещества, которые служат источником энергии, а также материалом для образования различных клеточных компонентов. От него зависит выведение из клетки токсических отходов, секреция различных полезных веществ и создание ионных градиентов, необходимых для нервной и мышечной активности, Изменение скорости переноса веществ может приводить к нарушениям биоэнергетических процессов, водно-солевого обмена, возбудимости и других процессов. Коррекция этих изменений лежит в основе действия многих лекарственных препаратов.
Существует два основных способа поступления веществ в клетку и вывода из клетки во внешнюю среду;
пассивный транспорт,
активный транспорт.
Пассивный транспорт идет по градиенту химической или электрохимической концентрации без затрат энергии АТФ. Если молекула транспортируемого вещества не имеет заряда, то направление пассивного транспорта определяется только разностью концентрации этого вещества по обеим сторонам мембраны (градиент химической концентрации). Если же молекула заряжена, то на ее транспорт влияют как градиент химической концентрации, так и электрический градиент (мембранный потенциал).
Оба градиента вместе составляют электрохимический градиент. Пассивный транспорт веществ может осуществляться двумя способами простой диффузией и облегченной диффузией.
При простой диффузии ионы солей и вода, могут проникать через селективные каналы. Эти каналы образуются за счет некоторых трансмембранных белков, формирующих сквозные транспортные пути, открытые постоянно или только на короткое время. Через селективные каналы проникают различные молекулы, имеющие соответствующие каналам размер и заряд.
Имеется и другой путь простой диффузии - это диффузия веществ через липидный бислой, через который легко проходят жирорастворимые вещества и вода. Липидный бислой непроницаем для заряженных молекул (ионов), и в то же время незаряженные малые молекулы могут свободно диффундировать, при этом, чем меньше молекула, тем быстрее она транспортируется. Довольно большая скорость диффузии воды через липидный бислой как раз и объясняется малой величиной ее молекул и отсутствием заряда.
При облегченной диффузии в транспорте веществ участвуют белки – переносчики, работающие по принципу «пинг-понг». Белок при этом существует в двух конформационных состояниях: в состоянии «понг» участки связывания транспортируемого вещества открыты с наружной стороны бислоя, а в состоянии «пинг» такие же участки открываются с другой стороны. Этот процесс обратимый. С какой же стороны в данный момент времени будет открыт участок связывания вещества, зависит от градиента концентрации, этого вещества.
Таким способом через мембрану проходят сахара и аминокислоты.
При облегченной диффузии скорость транспортировки веществ значительно возрастает в сравнении с простой диффузией.
Кроме белков-переносчиков, в облегченной диффузии принимают участие некоторые антибиотики, например, грамицидин и валиномицин.
Поскольку они обеспечивают транспорт ионов, их называют ионофорами .
Активный транспорт веществ в клетке. Этот вид транспорта всегда идет с затратой энергии. Источником энергии, необходимой для активного транспорта, является АТФ. Характерной особенностью этого вида транспорта является то, что он осуществляется двумя способами:
с помощью ферментов, называемых АТФ-азами;
транспорт в мембранной упаковке (эндоцитоз).
В наружной клеточной мембране присутствуют такие белки-ферменты, как АТФ-азы, функция которых заключается в обеспечении активного транспорта ионов против градиента концентрации. Поскольку они обеспечивают транспорт ионов, то этот процесс называют ионным насосом.
Известны четыре основные системы транспорта ионов в животной клетке. Три из них обеспечивают перенос через биологические мембраны.Na+ и К + , Са + , Н + , а четвертый - перенос протонов при работе дыхательной цепи митохондрии.
Примером механизма активного транспорта ионов может служить натрий-калиевый насос в животных клетках. Он поддерживает в клетке постоянную концентрацию ионов натрия и калия, которая отличается от концентрации этих веществ в окружающей среде: в норме в клетке ионов натрия бывает меньше, чем в окружающей среде, а калия - больше.
Вследствие этого по законам простой диффузии калий стремится уйти из клетки, а натрий диффундирует в клетку. В противовес простой диффузии натрий - калиевый насос постоянно выкачивает из клетки натрий и вводит калий: на три молекулы выбрасываемого наружу натрия приходится две молекулы вводимого в клетку калия.
Обеспечивает этот транспорт ионов натрий-калий зависимая АТФ-аза -фермент локализующийся в мембране таким образом, что пронизывает всю ее толщу, С внутренней стороны мембраны к этому ферменту поступает натрий и АТФ, а с наружной - калий.
Перенос натрия и калия через мембрану совершается в результате конформационных изменений, которые претерпевает натрий-калий зависимая АТФ-аза, активизирующаяся при повышении концентрации натрия внутри клетки или калия в окружающей среде.
Для энергообеспечения этого насоса необходим гидролиз АТФ. Этот процесс обеспечивает все тот же фермент натрий-калий зависимая АТФ-аза. При этом более одной трети АТФ, потребляемой животной клеткой в состоянии покоя, расходуется на работу натрий - калиевого насоса.
Нарушение правильной работы натрий - калиевого насоса приводит к различным серьезным заболеваниям.
КПД этого насоса превышает 50%, чего не достигают самые совершенные машины, созданные человеком.
Многие системы активного транспорта приводятся в действие за счет энергии, запасенной в ионных градиентах, а не путем прямого гидролиза АТФ. Все они работают как котранспортные системы (способствующие транспорту низкомолекулярных соединений). Например, активный транспорт некоторых сахаров и аминокислот внутрь животных клеток обусловливается градиентом иона натрия, причем чем выше градиент ионов натрия, тем больше скорость всасывания глюкозы. И, наоборот, если концентрация натрия в межклеточном пространстве заметно уменьшается, транспорт глюкозы останавливается. При этом натрий должен присоединиться к натрий - зависимому белку-переносчику глюкозы, который имеет два участка связывания: один для глюкозы, другой для натрия. Ионы натрия, проникающие в клетку, способствуют введению в клетку и белка-переносчика вместе с глюкозой. Ионы натрия, проникшие в клетку вместе с глюкозой, выкачиваются обратно натрий -калий зависимой АТФ-азой, которая, поддерживая градиент концентрации натрия, косвенным путем контролирует транспорт глюкозы.
Транспорт веществ в мембранной упаковке. Крупные молекулы биополимеров практически не могут проникать через плазмалемму ни одним из вышеописанных механизмов транспорта веществ в клетку. Они захватываются клеткой и поглощаются в мембранной упаковке, что получило название эндоцитоза . Последний формально разделяют на фагоцитоз и пиноцитоз. Захват клеткой твердых частиц - это фагоцитоз , а жидких - пиноцитоз . При эндоцитозе наблюдаются следующие стадии:
рецепция поглощаемого вещества за счет рецепторов в мембране клеток;
инвагинация мембраны с образованием пузырька (везикулы);
отрыв эндоцитозного пузырька от мембраны с затратой энергии – образование фагосомы и восстановление целостности мембраны;
Слияние фагосомы с лизосомой и образование фаголизосомы (пищеварительной вакуоли ) в которой происходит переваривание поглощенных частиц;
выведение непереваренного в фаголизосоме материала из клетки (экзоцитоз ).
В животном мире эндоцитоз является характерным способом питания многих одноклеточных организмов (например, у амеб), а среди много клеточных этот вид переваривания пищевых частиц встречается в энтодермальных клетках у кишечнополостных. Что касается млекопитающих и человека, то у них имеется ретикуло-гистио-эндотелиальная система клеток, обладающих способностью к эндоцитозу. Примером могут служить лейкоциты крови и купферовские клетки печени. Последние выстилают так называемые синусоидные капилляры печени и захватывают взвешенные в крови различные чужеродные частицы. Экзоцитоз - это и способ выведения из клетки многоклеточного организма секретируемого ею субстрата, необходимого для функции других клеток, тканей и органов.
Клеточные мембраны
В основе структурной организации клетки лежит мембранный принцип строения, то есть клетка в основном построена из мембран. Все биологические мембраны имеют общие структурные особенности и свойства.
В настоящее время общепринята жидкостно-мозаичная модель строения мембраны.
| Химический состав и строение мембраны |
Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами . Липиды составляют в среднем ≈40% химического состава мембраны. В бислое хвосты молекул в мембране обращены друг к другу, а полярные головки - наружу, поэтому поверхность мембраны гидрофильна. Липиды определяют основные свойства мембран.
Помимо липидов в состав мембраны входят белки (в среднем ≈60%). Они определяют большинство специфических функций мембраны. Молекулы белков не образуют сплошного слоя (рис. 280). В зависимости от локализации в мембране различают:
© периферические белки - белки, располагающиеся на наружной или внутренней поверхности липидного бислоя;
© полуинтегральные белки - белки, погруженные в липидный бислой на различную глубину;
© интегральные , или трансмембранные белки - белки, пронизывающие мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки.
Мембранные белки могут выполнять различные функции:
© транспорт определенных молекул;
© катализ реакций, происходящих на мембранах;
© поддержание структуры мембран;
© получение и преобразование сигналов из окружающей среды.
В состав мембраны может входить от 2 до 10% углеводов. Углеводный компонент мембран обычно представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Функции углеводов клеточной мембраны до конца не выяснены, однако можно сказать, что они обеспечивают рецепторные функции мембраны.
В животных клетках гликопротеины образуют надмембранный комплекс - гликокаликс , имеющий толщину в несколько десятков нанометров. В нем происходит внеклеточное пищеварение, располагаются многие рецепторы клетки, с его помощью, по-видимому, происходит адгезия клеток.
Молекулы белков и липидов подвижны, способны перемещаться, главным образом, в плоскости мембраны. Мембраны асимметричны, то есть липидный и белковый состав наружной и внутренней поверхности мембраны различен.
Толщина плазматической мембраны в среднем 7,5 нм.
Одна из основных функций мембраны - транспортная, обеспечивающая обмен веществ между клеткой и внешней средой. Мембраны обладают свойством избирательной проницаемости, то есть хорошо проницаемы для одних вещества или молекул и плохо проницаемы (или совсем непроницаемы) для других. Проницаемость мембран для разных веществ зависит и от свойств их молекул (полярность, размер и т.д.), и от характеристики мембран (внутренняя часть липидного слоя гидрофобна).
Существуют различные механизмы транспорта веществ через мембрану (рис. 281). В зависимости от необходимости использования энергии для осуществления транспорта веществ, различают:
© пассивный транспорт - транспорт веществ, идущий без затрат энергии;
© активный транспорт - транспорт, идущий с затратами энергии.
| Пассивный транспорт |
В основе пассивного транспорта лежит разность концентраций и зарядов. При пассивном транспорте вещества всегда перемещаются из области с более высокой концентрацией в область с более низкой, то есть по градиенту концентрации. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Различают три основных механизма пассивного транспорта:
© Простая диффузия - транспорт веществ непосредственно через липидный бислой. Через него легко проходят газы, неполярные или малые незаряженные полярные молекулы. Чем меньше молекула и чем более она жирорастворима, тем быстрее она проникает через мембрану. Интересно, что вода, несмотря на то, что она относительно нерастворима в жирах, очень быстро проникает через липидный бислой. Это объясняется тем, что ее молекула мала и электрически нейтральна. Диффузию воды через мембраны называют осмосом .
Диффузия через мембранные каналы . Заряженные молекулы и ионы (Na + , K + , Ca 2+ , Cl -) не способны проходить через липидный бислой путем простой диффузии, тем не менее, они проникают через мембрану, благодаря наличию в ней особых каналообразующих белков, формирующих водяные поры.© Облегченная диффузия - транспорт веществ с помощью специальных
транспортных белков, каждый из которых отвечает за транспорт определенных молекул или групп родственных молекул. Они взаимодействуют с молекулой переносимого вещества и каким-либо способом перемещают ее сквозь мембрану. Таким образом в клетку транспортируются сахара, аминокислоты, нуклеотиды и многие другие полярные молекулы.
| Активный транспорт |
Необходимость активного транспорта возникает тогда, когда требуется обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ.
Одной из наиболее изученных систем активного транспорта является натрий-калиевый насос. Концентрация K внутри клетки значительно выше, чем за ее пределами, а Na - наоборот. Поэтому К через водяные поры мембраны пассивно диффундирует из клетки, а Na - в клетку. Вместе с тем, для нормального функционирования клетке важно поддерживать определенное соотношение ионов К и Na в цитоплазме и во внешней среде. Это оказывается возможным потому, что мембрана, благодаря наличию (Na + K)-насоса, активно перекачивает Na из клетки, а K в клетку. На работу (Na + K)-насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки.
Насос представляет собой особый трансмембранный белок мембраны, способный к конформационным изменениям, благодаря чему он может присоединять к себе как ионы К, так и ионы Na. Цикл работы (Na + K)-насоса складывается из нескольких фаз (рис. 282):
© с внутренней стороны мембраны к белку-насосу поступают ионы Na и молекула АТФ, а с наружной - ионы K;
© ионы Na соединяются с молекулой белка, и белок приобретает АТФ-азную активность, то есть приобретает способность вызывать гидролиз АТФ, сопровождающийся освобождением энергии, приводящей в движение насос;
© освободившийся при гидролизе АТФ фосфат присоединяется к белку, то есть происходит фосфорилирование белка;
© фосфорилирование вызывает конформационные изменения белка, он оказывается неспособным удерживать ионы Na - они высвобождаются и выходят за пределы клетки;
© новая конформация белка такова, что оказывается возможным присоединение к нему ионов K;
© присоединение ионов K вызывает дефосфорилирование белка, в результате чего он вновь изменяет свою конформацию;
© изменение конформации белка приводит к высвобождению ионов K внутри клетки;
© теперь белок вновь готов присоединить к себе ионы Na.
За один цикл работы насос выкачивает из клетки 3 иона Na и закачивает 2 иона К. Такая разница в количестве переносимых ионов связана с тем, что проницаемость мембраны для ионов К выше, чем для ионов Na. Соответственно K быстрее пассивно диффундирует из клетки, чем Na в клетку.
крупных частиц (например, фагоцитоз лимфоцитов, простейших и др.);© пиноцитоз - процесс захвата и поглощения капелек жидкости с растворенными в ней веществами.
Экзоцитоз - процесс выведения различных веществ из клетки. При экзоцитозе мембрана везикулы (или вакуоли), при соприкосновении с наружной цитоплазматической мембраной, сливается с ней. Содержимое везикулы выводится за пределы летки, а ее мембрана включается в состав наружной цитоплазматической мембраны.


